Any suggestions as to alternate etymologies are welcome. Based on my understanding of the Thai language, and my studies of other languages, and what I can find online, here's where I think these 10 words may have come from:
สิงโต [siŋtɔ:] (n. lion), first element cognate to Hindi "सिंह" [siŋ] (lion), from Sanskrit [siṃha] (lion), cognate to Swahili "simba" (lion).
ตา [ta:] (n. eye), possibly shortened from Proto-Austronesian *mata (eye), as in Malay "mata" (eye) and Tagalog "mata" (eye), likely sharing a common origin with Proto-Austro-Asiatic *mat (eye), as in Vietnamese "mắt" (eye).
สาม [sa:m] (three), of Sinitic origin, cognate to Chinese "三" [sam] (three) and Japanese "三" [san] (three).
ดำ [da:m] (black), possible cognate to Proto-Vietic *tem (night) and Proto-Monic *tam (night).
นก [nuk] (n. bird), possibly shortened from Proto-Austronesian *manuk (chicken). Probably cognate to White Hmong "noog" [noŋ] (bird) as well.
สวัสดี [sàwàtdiː] (hello, goodbye), of Indo-European origin, from Sanskrit "स्वस्ति" [svasti] (well-being, luck), cognate to English "swastika" (Greek cross with arms bent at right angles, originally a good luck charm). Svasti = [su] (good) + [asti] (is), second element from Proto-Indo-European *es-ti (is), cognate to English "is" (is) and German "ist" (is).
โรง [rɔ:ŋ] (n. building, structure), borrowed from Old Khmer [roŋ] (shelter/room/hall), c.f. Khmer [raoŋ], from Proto-Austro-Asiatic *ro:ŋ (hall/house).
ข้าว [ka:w] (n. rice), cognate to Proto-Austro-Asiatic *kɔw (rice), cognate to Vietnamese "gạo" (rice). Direction of borrowing between the language families is uncertain, and probably took place many thousands of years ago.
ไฟ [fai] (electric), possibly of recent English origin, possibly cognate to English "fire" (fire), from Proto-Indo-European *paewr (fire). First element of Lao [fai-fa:] is clearly a cognate, and White Hmong "fai fab" [fai fa] was clearly borrowed from the Lao form.
แอปเปิล [ɛːpɤːl] (n. apple), of recent English origin, cognate to Malay "epal" (apple) and English "apple" (apple), from Proto-Indo-European *abel (apple).
เปล่า [plaɔ] (not, blank, empty, nil), possibly cognate to Lao "ບໍ່" [bɔː] (not).
Wednesday, November 16, 2011
Sunday, November 13, 2011
Genetic Evidence Regarding the Peopling of Siberia
In modern times, the expanse of Siberia is dominated by speakers of Slavic languages (including Russian and Ukranian), Altaic languages (including languages of the Mongolic, Turkic and Tungusic language families) and Uralic languages (including languages of the Samoyedic language family). However, there are several language families and language isolates found throughout Siberia that tell of a more distant history of the population of Siberia. These languages include the Chukotko-Kamchatkan languages of eastern Siberia, the Yukaghir languages of western Siberia, the Nivkh and Ainu language isolates of the Sakhalin, and the Ket language (the only surviving language of the Yeniseian language family).
Additionally, based on the theory that the Americas were populated by Siberians who crossed the Bering Strait, one would expect that languages ancestral to the Eskimo-Aleut, Na-Dené and Amerind language families of the indigenous Americans must have at one time been spoken in Siberia. In 2008, historical linguist Edward Vajda proposed that the Na-Dené and Yeniseian language families are in fact subphyla of a larger Dené-Yeniseian language stock, a theory which has since received wide acceptance. A similar proposal by Michael Fortescue in 1998 suggests that the Uralic, Chukotko-Kamchatkan, Yukaghir and Eskimo-Aleut families are in fact branches of a larger Uralo-Siberian language family.
This article explores the genetic differences and similarities between the modern speakers of the Slavic, Uralo-Siberian, Altaic, Nivkh, Ainu, Dené-Yeniseian and Amerind language families, with the goal of understanding how, when and why their ancestors came to inhabit the harsh expanse of Siberia.
The Big Picture: Haplogroup K(xLT) vs. Haplogroup C
In terms of Y-DNA haplogroups exhibited by modern inhabitants of Siberia, subclades of haplogroup C and of haplogroup K(xLT) dominate the region. These two distantly related haplogroups (sharing a most recent common ancestor 60,000 years before present) are believed to have spread across Eurasia independently, although they ultimately met in Siberia, forming the basis of the current population of both Siberia and the Americas. A lack of haplogroup C among the speakers of Amerind languages suggests either a bottleneck in the migration through the Americas, or that ancestors of the first Siberians to migrate to the Americas lived in an eastern Siberian region that had thus far been populated by K(xLT), but not C ethnicities. I find the latter to be a more convincing theory, and propose that an examination of the peopling of Siberia should begin with an examination of the ratio of C to K(xLT) among present populations.
Most Recent Arrivals: Slavic Speakers
The spread of Slavic languages throughout Siberia is known to be recent. Based on linguistic research alone, the Slavic languages probably only split off from the Indo-European language family less than 4,000 years ago, and most Slavic languages are not spoken in Siberia, but rather Eastern Europe. Speakers of Slavic languages exhibit a high occurrence of Y-DNA haplogroup R1a, associated with Indo-European languages, a sister clade of R1b which is associated with the Vasconian languages of the Iberian Peninsula. The Slavic people do not generally exhibit haplogroup C. Any population speaking a Slavic language in Siberia would either be the result of recent migration (as would be indicated by the dominance of R1a, or recent adoption of the Slavic language by that population, i.e. language shift. It should be noted, however, that R1a is a clade of K(xLT), indicating a distant relation to some of the other Siberian ethnicities, albeit some 40,000 years before present, and through a most recent common ancestor who probably resided in Central Asia, not Siberia.
Populations With Large Percentages of Haplogroup C
According to a 2004 study by Kristiina Tambet, et al., speakers of Tungusic languages (Evens and Evenks, in this study) exhibited high proportions of haplogroup C. Evens exhibited 74.2% haplogroup C; Evenks exhibited 67.7%. The dominant K(xLT) haplogroup making up the remainder of these populations was haplogroup N, perhaps a result of recent or ancient admixture with Uralo-Siberian speaking populations. A 2005 study by Miroslava Derenko demonstrated that Mongolic speaking Kalmyks exhibited 70.6% haplogroup C; Buryats exhibited 63.9% haplogroup C; and Mongolians exhibited 53.8%. In each of these populations, the next largest component of the Y-DNA mixture consisted of haplogroup N (although Mongolians exhibited a similar proportion of haplogroup O, likely due to a long period of close contact with Chinese populations). Due to the time-depth of the separation of the Tungusic and Mongolic languages from the Altaic family, the haplogroup N component probably represents a uniform superstrata that manifested in Altaic populations soon after Altaic arrival in Siberia. While Turkic speaking Siberian populations exhibit both haplgroups C and N, they exhibit haplogroup C in far lesser proportions, probably due to admixture from various populations as a result of centuries of migrations and reverse migrations along the silk road. The dominant clade of haplogroup C among Altaic speakers is C3, which is believed to have first occurred approximately 12,000 years ago. C3 is also present in over 11% of the hypothetically Macro-Altaic speakers of the Korean language, and the related C1 sister clade, also approximately 12,000 years old, is frequent in hypothetically Macro-Altaic Japanese speakers. Perhaps the Macro-Altaic languages of the Japanese and Korean populations split off from the rest of the Altaic languages very close to the time subclades C1 and C3 occurred. Other Y-DNA admixtures in Japanese and Koreans are not discussed here, as they are not strictly-speaking, Siberian populations. The Ainus of Siberia and Japan, however, will be discussed in a subsequent section of this article. It suffices to say that populations of Proto-Macro-Altaic speakers arrived in Siberia, probably in two migrations (C1 along the coast and C3 further inland), around 10,000 years ago, and soon after, the inland population began interbreeding with earlier Uralo-Siberian populations exhibiting haplogroup N.
Subdivisions of Haplogroup K(xLT)
At the timeframe of around 40,000 years before present, K(xLT) was probably already divided into four subclades, namely M, NO, P and S. Haplogroups M and S are not of particular importance to this discussion, as they generally appear only in island populations of the southern Pacific, such as New Guinea. Approximately 30,000 years ago, haplogroup O split off from NO, and descendants of the individual who first carried that mutation include much of the population of central and southeast Asia (including Sino-Tibetan, Tai-Kadai, Austronesian, Hmong-Mien and Austro-Asiatic speakers). Haplogroup N probably first occurred around 25,000 years before present, and its N1 subclades (primarily N1c) probably arrived in Siberia around 15,000 years ago, and spread across the region via Uralo-Siberian speaking populations. Haplogroup Q (the predominant Siberian subclade of haplogroup P) probably predated haplgroup N in Siberia, and will be discussed subsequently.
Evidence strongly suggests that the progenitor of haplogroup N spoke a language ancestral to the Uralic languages, as haplogroup N is found in high proportions in all areas where Uralic languages are spoken, particularly in populations of Nenets (97.3%), Nganasans (92.1%) and Khants (76.6%). See also, the Tambet study, cited above. Selkups appear to be outliers, exhibiting only 6.9% haplogroup N, but 66.4% haplogroup Q, indicating a likely language shift of a Q population that assimilated into Uralic culture. According to a 2001 study by Jeffrey Lell, et al., 50.6% of Yupiks (Eskimo-Aleut speakers) are also of haplogroup N, as are 58.3% of Chukchis (Chukotko-Kamchatkan speaker), which supports the Uralo-Siberian hypothesis. I was not able to find any statistics regarding the levels of N among Yukaghir speakers. Although Koryaks speak a Chukotko-Kamchatkan language, they only exhibit 22.2% haplogroup N, and appear to have interbred heavily with Altaic speaking populations. The same is true regarding Itelmens, who only exhibit 11% haplogroup N, and have large components of both Altaic and Slavic admixture (C3 and R1a, respectively).
Origin of the Ainu
It would appear that the Ainu may be remnants of the first migration from Africa to Asia, as they exhibit haplogroup D (specifically D2), which is a sister clade of haplogroup E (common among Nilo-Saharan populations in Africa). Ainu also exhibit a small percentage of haplogroup C3, suggesting admixture from an unrelated population in Siberia, probably the Nivkhs who co-habited with the Ainu on Sakhalin Island prior to the Ainu migration to Japan, and who exhibit exhibit C3 at a rate of 47%. If the Ainu language was found to have any recognizable relation to any of the other Siberian languages, it would have to be a result of language shift, as the Ainu are not believed to be closely related to any other Siberian population, with the exception of the Nivkh admixture mentioned above. While the Ainu are the only population in Siberia with a strong correspondence to haplogroup D, other populations with considerable proportions of haplogroup D include Negrito populations in southern Asia, especially aboriginal Andaman Islanders. Perhaps mt-DNA analysis can explain the difference in appearance between the Ainu and Negrito populations. If the founding population of the Ainu consisted of mostly males, and if those males interbred early-on with females from another Siberian population (such as the Nivkhs), then one would expect to see haplogroup D in terms of Y-DNA, but mostly common Siberian mt-DNA haplogroups.
Haplogroup Q
With the possible exception of the paternal ancestors of the Ainu, it would appear that the earliest settlers of Siberia were populations exhibiting haplogroup Q. In fact, haplogroup Q underlies all of the previously discussed populations in low frequencies (possibly excepting the Ainus, for which I have not found any data relating to haplogroup Q frequency). This leaves us with one haplogroup with which to explain the divergence of the Nivkh, Dené-Yeniseian and Amerind languages, all exhibiting high proportions of haplogroup Q. Lell's study puts the Nivkh population at 35% Q. Kets (Dené-Yeniseian) have a 93.7% incidence of Q according to Tambet.
Michael Fortescue has suggested that the Nivkh language may be connected to a Native American language family referred to by Edward Sapir in 1929 as the Mosan language family (a subgroup of the traditional Amerind grouping that includes Salishan, Wakashan, and Chimakuan languages of the Pacific Northwest). According to Wikipedia, however, it has not yet even been proven whether the Mosan language family is in fact monophylectic. I would suggest, that based on Y-DNA evidence, a Paleosiberian language ancestral to the Nivkh, Dené-Yeniseian and Amerind languages, was probably once spoken in eastern Siberia, prior to the arrival of the Uralo-Siberian, Altaic and Slavic speakers. I am not suggesting at this point that Amerind is a monophylectic group, and in fact there seems to be some evidence that some Amerind languages are more closely related to Nivkh than they are to other Amerind languages. The key to reconstructing a Paleosiberian proto-language will be first studying, classifying and reconstructing the proto-languages of language families of the Americas, and in particular Joseph Greenburg's illusive Amerind languages.
Additionally, based on the theory that the Americas were populated by Siberians who crossed the Bering Strait, one would expect that languages ancestral to the Eskimo-Aleut, Na-Dené and Amerind language families of the indigenous Americans must have at one time been spoken in Siberia. In 2008, historical linguist Edward Vajda proposed that the Na-Dené and Yeniseian language families are in fact subphyla of a larger Dené-Yeniseian language stock, a theory which has since received wide acceptance. A similar proposal by Michael Fortescue in 1998 suggests that the Uralic, Chukotko-Kamchatkan, Yukaghir and Eskimo-Aleut families are in fact branches of a larger Uralo-Siberian language family.
This article explores the genetic differences and similarities between the modern speakers of the Slavic, Uralo-Siberian, Altaic, Nivkh, Ainu, Dené-Yeniseian and Amerind language families, with the goal of understanding how, when and why their ancestors came to inhabit the harsh expanse of Siberia.
The Big Picture: Haplogroup K(xLT) vs. Haplogroup C
In terms of Y-DNA haplogroups exhibited by modern inhabitants of Siberia, subclades of haplogroup C and of haplogroup K(xLT) dominate the region. These two distantly related haplogroups (sharing a most recent common ancestor 60,000 years before present) are believed to have spread across Eurasia independently, although they ultimately met in Siberia, forming the basis of the current population of both Siberia and the Americas. A lack of haplogroup C among the speakers of Amerind languages suggests either a bottleneck in the migration through the Americas, or that ancestors of the first Siberians to migrate to the Americas lived in an eastern Siberian region that had thus far been populated by K(xLT), but not C ethnicities. I find the latter to be a more convincing theory, and propose that an examination of the peopling of Siberia should begin with an examination of the ratio of C to K(xLT) among present populations.
Most Recent Arrivals: Slavic Speakers
The spread of Slavic languages throughout Siberia is known to be recent. Based on linguistic research alone, the Slavic languages probably only split off from the Indo-European language family less than 4,000 years ago, and most Slavic languages are not spoken in Siberia, but rather Eastern Europe. Speakers of Slavic languages exhibit a high occurrence of Y-DNA haplogroup R1a, associated with Indo-European languages, a sister clade of R1b which is associated with the Vasconian languages of the Iberian Peninsula. The Slavic people do not generally exhibit haplogroup C. Any population speaking a Slavic language in Siberia would either be the result of recent migration (as would be indicated by the dominance of R1a, or recent adoption of the Slavic language by that population, i.e. language shift. It should be noted, however, that R1a is a clade of K(xLT), indicating a distant relation to some of the other Siberian ethnicities, albeit some 40,000 years before present, and through a most recent common ancestor who probably resided in Central Asia, not Siberia.
Populations With Large Percentages of Haplogroup C
According to a 2004 study by Kristiina Tambet, et al., speakers of Tungusic languages (Evens and Evenks, in this study) exhibited high proportions of haplogroup C. Evens exhibited 74.2% haplogroup C; Evenks exhibited 67.7%. The dominant K(xLT) haplogroup making up the remainder of these populations was haplogroup N, perhaps a result of recent or ancient admixture with Uralo-Siberian speaking populations. A 2005 study by Miroslava Derenko demonstrated that Mongolic speaking Kalmyks exhibited 70.6% haplogroup C; Buryats exhibited 63.9% haplogroup C; and Mongolians exhibited 53.8%. In each of these populations, the next largest component of the Y-DNA mixture consisted of haplogroup N (although Mongolians exhibited a similar proportion of haplogroup O, likely due to a long period of close contact with Chinese populations). Due to the time-depth of the separation of the Tungusic and Mongolic languages from the Altaic family, the haplogroup N component probably represents a uniform superstrata that manifested in Altaic populations soon after Altaic arrival in Siberia. While Turkic speaking Siberian populations exhibit both haplgroups C and N, they exhibit haplogroup C in far lesser proportions, probably due to admixture from various populations as a result of centuries of migrations and reverse migrations along the silk road. The dominant clade of haplogroup C among Altaic speakers is C3, which is believed to have first occurred approximately 12,000 years ago. C3 is also present in over 11% of the hypothetically Macro-Altaic speakers of the Korean language, and the related C1 sister clade, also approximately 12,000 years old, is frequent in hypothetically Macro-Altaic Japanese speakers. Perhaps the Macro-Altaic languages of the Japanese and Korean populations split off from the rest of the Altaic languages very close to the time subclades C1 and C3 occurred. Other Y-DNA admixtures in Japanese and Koreans are not discussed here, as they are not strictly-speaking, Siberian populations. The Ainus of Siberia and Japan, however, will be discussed in a subsequent section of this article. It suffices to say that populations of Proto-Macro-Altaic speakers arrived in Siberia, probably in two migrations (C1 along the coast and C3 further inland), around 10,000 years ago, and soon after, the inland population began interbreeding with earlier Uralo-Siberian populations exhibiting haplogroup N.
Subdivisions of Haplogroup K(xLT)
At the timeframe of around 40,000 years before present, K(xLT) was probably already divided into four subclades, namely M, NO, P and S. Haplogroups M and S are not of particular importance to this discussion, as they generally appear only in island populations of the southern Pacific, such as New Guinea. Approximately 30,000 years ago, haplogroup O split off from NO, and descendants of the individual who first carried that mutation include much of the population of central and southeast Asia (including Sino-Tibetan, Tai-Kadai, Austronesian, Hmong-Mien and Austro-Asiatic speakers). Haplogroup N probably first occurred around 25,000 years before present, and its N1 subclades (primarily N1c) probably arrived in Siberia around 15,000 years ago, and spread across the region via Uralo-Siberian speaking populations. Haplogroup Q (the predominant Siberian subclade of haplogroup P) probably predated haplgroup N in Siberia, and will be discussed subsequently.
Evidence strongly suggests that the progenitor of haplogroup N spoke a language ancestral to the Uralic languages, as haplogroup N is found in high proportions in all areas where Uralic languages are spoken, particularly in populations of Nenets (97.3%), Nganasans (92.1%) and Khants (76.6%). See also, the Tambet study, cited above. Selkups appear to be outliers, exhibiting only 6.9% haplogroup N, but 66.4% haplogroup Q, indicating a likely language shift of a Q population that assimilated into Uralic culture. According to a 2001 study by Jeffrey Lell, et al., 50.6% of Yupiks (Eskimo-Aleut speakers) are also of haplogroup N, as are 58.3% of Chukchis (Chukotko-Kamchatkan speaker), which supports the Uralo-Siberian hypothesis. I was not able to find any statistics regarding the levels of N among Yukaghir speakers. Although Koryaks speak a Chukotko-Kamchatkan language, they only exhibit 22.2% haplogroup N, and appear to have interbred heavily with Altaic speaking populations. The same is true regarding Itelmens, who only exhibit 11% haplogroup N, and have large components of both Altaic and Slavic admixture (C3 and R1a, respectively).
Origin of the Ainu
It would appear that the Ainu may be remnants of the first migration from Africa to Asia, as they exhibit haplogroup D (specifically D2), which is a sister clade of haplogroup E (common among Nilo-Saharan populations in Africa). Ainu also exhibit a small percentage of haplogroup C3, suggesting admixture from an unrelated population in Siberia, probably the Nivkhs who co-habited with the Ainu on Sakhalin Island prior to the Ainu migration to Japan, and who exhibit exhibit C3 at a rate of 47%. If the Ainu language was found to have any recognizable relation to any of the other Siberian languages, it would have to be a result of language shift, as the Ainu are not believed to be closely related to any other Siberian population, with the exception of the Nivkh admixture mentioned above. While the Ainu are the only population in Siberia with a strong correspondence to haplogroup D, other populations with considerable proportions of haplogroup D include Negrito populations in southern Asia, especially aboriginal Andaman Islanders. Perhaps mt-DNA analysis can explain the difference in appearance between the Ainu and Negrito populations. If the founding population of the Ainu consisted of mostly males, and if those males interbred early-on with females from another Siberian population (such as the Nivkhs), then one would expect to see haplogroup D in terms of Y-DNA, but mostly common Siberian mt-DNA haplogroups.
Haplogroup Q
With the possible exception of the paternal ancestors of the Ainu, it would appear that the earliest settlers of Siberia were populations exhibiting haplogroup Q. In fact, haplogroup Q underlies all of the previously discussed populations in low frequencies (possibly excepting the Ainus, for which I have not found any data relating to haplogroup Q frequency). This leaves us with one haplogroup with which to explain the divergence of the Nivkh, Dené-Yeniseian and Amerind languages, all exhibiting high proportions of haplogroup Q. Lell's study puts the Nivkh population at 35% Q. Kets (Dené-Yeniseian) have a 93.7% incidence of Q according to Tambet.
Michael Fortescue has suggested that the Nivkh language may be connected to a Native American language family referred to by Edward Sapir in 1929 as the Mosan language family (a subgroup of the traditional Amerind grouping that includes Salishan, Wakashan, and Chimakuan languages of the Pacific Northwest). According to Wikipedia, however, it has not yet even been proven whether the Mosan language family is in fact monophylectic. I would suggest, that based on Y-DNA evidence, a Paleosiberian language ancestral to the Nivkh, Dené-Yeniseian and Amerind languages, was probably once spoken in eastern Siberia, prior to the arrival of the Uralo-Siberian, Altaic and Slavic speakers. I am not suggesting at this point that Amerind is a monophylectic group, and in fact there seems to be some evidence that some Amerind languages are more closely related to Nivkh than they are to other Amerind languages. The key to reconstructing a Paleosiberian proto-language will be first studying, classifying and reconstructing the proto-languages of language families of the Americas, and in particular Joseph Greenburg's illusive Amerind languages.
Wednesday, November 9, 2011
Classification of the Minoan Language
A portion of Andrew Oh-Willeke's analysis in his recent blog post entitled Notes On Ancient European Y-DNA and Ancient European Cultures caught my attention:
While researching the subject, I came upon a linguistics doctoral thesis entitled The Structure of the Minoan Language by Gareth Owens of the National and Kapodistrian University of Athens, which proposes that the Minoan language was in fact an Indo-European language, and likely belongs to a separate branch of the language family than any of the living Indo-European languages. I found his reasoning to be quite convincing, as it is based not only on lexical similarities that could be explained by borrowing, but also on morphological similarities, including masculine, feminine and neuter noun declension patterns. Owens had studied the Minoan Language for approximately 15 years at the time this paper was written (2004), and stated:
The Minoans had a script that probably recounted a language rather than a proto-language in a somewhat phonetic way, but it has not been deciphered and the extant Minoan Linear A script appears to consist mostly of bureaucratic records of ration programs, taxes and religious sacrifices. My own impression is that Minoan bore a striking similarity to the non-Indo-European language of the Hattic people whose language was superceded by the Indo-European Hittites in the time period from ca. 2000 BCE to 1500 BCE, by which time Hattic had ceased to be used outside religious ceremonies.I found this interesting for two reasons. First off, I having only researched living languages in connection with my DNA articles thus far, I wasn't aware of this extinct, unclassified, potentially non-Indo-European language attested in Europe (on the island of Crete). Secondly, the idea that it "has not been deciphered" sounded intriguing.
While researching the subject, I came upon a linguistics doctoral thesis entitled The Structure of the Minoan Language by Gareth Owens of the National and Kapodistrian University of Athens, which proposes that the Minoan language was in fact an Indo-European language, and likely belongs to a separate branch of the language family than any of the living Indo-European languages. I found his reasoning to be quite convincing, as it is based not only on lexical similarities that could be explained by borrowing, but also on morphological similarities, including masculine, feminine and neuter noun declension patterns. Owens had studied the Minoan Language for approximately 15 years at the time this paper was written (2004), and stated:
It has now been possible to discuss and offer an interpretation, both etymologically and morphologically, for 50 words that constitute the Lexicon of the Minoan language. Linear A inscriptions, when read with Linear B sound values, and when interpreted in the Minoan cultural context, make sense as an Indo-European language of the Second Millennium B.C. . . I would place the Minoan language between Greek and Armenian as a distinct branch of the Indo-European family of languages, with whatever caveat we must keep in mind of what surprises scientific research may have for us in the future.With respect to the Y-DNA haplogroups of the Minoan speakers, according to an article entitled Middle Eastern and European mtDNA lineages characterize populations from eastern Crete by Laisel Martinez, Sheyla Mirabal, Javier R. Luis and Rene J. Herrera, published in the American Journal of Physical Anthropology, as quoted in Mathilda's Anthropology Blog:
Haplogroups J2a1h-M319 (8.8%) and J2a1b1-M92 (2.6%) [among the current population on Crete] were associated with the Minoan culture linked to a late Neolithic/Early Bronze Age migration to Crete ca. 3100 BCE from North-Western/Western Anatolia and Syro-Palestine (ancient Canaan, Levant, and pre-Akkadian Anatolia); Aegean prehistorians link the date 3100 BCE to the origins of the Minoan culture on Crete.Interestingly, y-DNA haplogroup J (common in the Caucasus, ancestral to both J2a1h and J2a1b1), along with haplogroup I (common where Germanic languages are spoken) are believed to correspond with the early Cro-Magnon settlement of Europe, while haplogroup R1a is commonly associated with the Indo-European expansion. Hopefully, further deciphering of the Minoan language inscriptions will shed light on the apparent divergence between the genetic and linguistic conclusions reached regarding the Minoan people.
Monday, November 7, 2011
New Evidence Relating to the Ancestry of Lars Magnus Winblad

Pictured is my great-great grandfather John Edward Winblad, a native of Sweden, son of Anton Julius Winblad, grandson of Lars Magnus Winblad. John was the maternal grandfather of my maternal grandmother Helen Eloise (Freudenberg) Borland-Brindley. While no direct paternal descendants of John have yet tested their y-DNA (most of his living descendants are either female or descend from his daughters' lines), today I stumbled upon a record that might provide some clues as to the origin of my Winblad family.
Some time ago, my cousin Richard Norton commissioned research on Anton Julius Winblad, our earliest known Winblad ancestor at the time, and a hired genealogist in Sweden revealed records indicating Anton was the son of a Lars Magnus Winblad, born June 3, 1794 or 1797 in Vingåker, Södermanland, Sweden. No birth or baptismal record has been found for Lars. A year or two ago, I found a user-submitted family tree on the LDS website FamilySearch that included an Erick Vingblad born December 12, 1798 in Vingåker married to a woman by the name of Britta Ehrsdatter. See here. Suspecting Lars and Erick were brothers, I searched for Erick's marriage record, and eventually (as more records have been added to the FamilySearch collection) found reference to the marriage record of Erick and Britta. See here. Erick is listed as "Eric Winbladh," and Britta as "Brita Ersdr." The marriage took place July 1, 1821 in Vastra (West) Vingåker.
Today, I spent several hours looking thorugh the online baptismal records of Vingåker for Lars' baptism, but to no avail. However, I found a new piece of circumstantial evidence that might lead to a major breakthrough in tracing back my Winblad family. The baptismal records use patronyms, not surnames, so there are no Winblads listed. However, I found that the only Eric(k) whose birth was recorded in Vingåker on December 12, 1798 was a child christened as "Eric Nilsdr." The record clearly states that Eric was a male, although his patronymic name indicates "daughter of Nils." Eric's father is not listed in the record, which is highly unusual, but his mother is listed as "Anna Nilsdr." Perhaps the father of Eric, and presumably Lars, either died or left the family prior to Eric's birth. The baptismal record for Eric can be accessed here, although a scanned image of the record is not yet available online.
Eric is a pretty common Swedish name, and so the likelihood of more than one Eric having been born in Vingåker on a given day should not be discounted where a surname cannot be confirmed. However, even today, the population of Vingåker is approximately 4300, after a considerable expansion in the mid-1800s due to Vingåker's location along a railroad line. Even assuming the population of Vingåker was 4300, 200 years before present, only 2150 would have been females, and far less than 1,000 of those females would have given birth in 1798. Statistically, there would have been less than three births per day, so the odds of more than one child named Eric having been born in Vingåker on December 12, 1798 is unlikely. However, there is no definitive proof that Lars and Eric were brothers, and to prove such a contention, y-DNA samples are needed from each line for examination. Also, this new piece of evidence only tells that Lars' mother might have been Anna Nilsdotter, and does not provide any other family information that could be used to trace the family back further. In and around Vingåker, for example, in the 20 to 40 years prior to the births of Lars and Eric, there are many marriages recorded for women named Anna who were daughters of fathers named Nils.
Sunday, November 6, 2011
Mt-DNA Matches
This past year, my cousin Richard Norton and I had our mitochondrial DNA tested for genealogical purposes. The results provide information about my direct maternal lineage, and about my father's direct maternal lineage (since Richard and my father were sons of two sisters, Selma and Helen Freudenberg). A brief analysis of the results of each test follows:
My Direct Maternal Line

Pictured is my great-grandmother Eva Rice. Eva is my earliest known direct maternal ancestor. She was born around October 31, 1883 in Lithuania. Her name at birth was Ieva Daukšaitė, shortened to Eva Douse after immigrating to the United States. She took the surname Rice after marrying my great-grandfather Peter Rice, a descendant of the Lithuanian Račius family. Eva's father was Jonas Daukša. Her mother, whose name is not currently known, died in Lithuania, probably giving birth to Eva. For a full biography of Eva Rice, see my previous article on Familypedia.
Based on a sample of my mitochondrial DNA, Eva was of mt-DNA haplogroup T2. My closest mt-DNA match (perfect match to my HVR1 and HVR2) is with an individual from Bulgaria, whose earliest known maternal ancestor had the Slavic maiden name Fiyodorova. My next two closest matches (perfect match to my HVR1, but they did not test their HVR2) are with individuals whose earliest known maternal ancestors are of Polish origin, with maiden names Borek and Stefineski. The common maternal ancestor of all four lineages was likely a woman who lived in Poland or the Ukrane less than 2000 years ago. Although Lithuanians speak a Baltic language, it is not surprising that a person from Lithuania would have Slavic, and particularly Polish ancestry. Based on y-chromosomal DNA studies of Baltic populations, 38% of Lithuanians are of y-DNA haplogroup R1a, as are 55% of the population of Poland. (Another 42% of Lithuanians are of y-DNA haplogroup N, indicating relationship to Uralic speakers.)
Mt-DNA haplogroup T (ancestral to T2) likely arrived in Poland long before Indo-European y-DNA haplogroup R1a, as the incidence of haplogroup T corresponds more directly with R1 in Europe, generally, than it does with either of its R1a or earlier arriving R1b subclades individually (which, for reasons beyond the scope of this article, does not imply that mt-DNA haplogroup T and y-DNA haplogroup R1 are necessarily closely related).
The individual woman who first had the mutation that defined mt-DNA haplogroup T, my direct maternal ancestor, has been given the nickname "Tara" by geneticists. Tara probably lived in the Near East, more than 45,000 years ago. Tara was also a direct maternal ancestor of outlaw Jesse James, and Tsar Nicholas II. The International Society of Genetic Genealogy places haplogroup T2 (shared by Jesse James, Tsar Nicholas II and myself) in Europe sometime between the late Stone Age and 8000 BC.
My Father's Direct Maternal Line

This is a photograph of my great-grandmother Maria Freudenberg (1895-1987), the common direct maternal ancestor of my father and my cousin Richard Norton who shared his mt-DNA results with me. Maria was born Maria Elizabeth Winblad. Maria's mother was Salmine Sofie Severine Olsdatter (1862-1914), who was born in Farsund, Vest-Agder, Norway. Salmine's mother was Thea Johanne Tostensdatter (1825-1865), born in Kristiansand, Vest-Agder, Norway. Thea's mother was Johanne Jacobsdatter (1795-?), born in Eigersund, Rogaland, Norway. Johanne's mother was Gitlaug Tollaksdatter (1773-1806), born in Hå, Rogaland, Norway. Gitlaug's mother was Ingeborg Pedersdatter (c1747-?), believed to have been born in Eigersund.
Based on a sample of my cousin's mitochondrial DNA, Ingeborg Pedersdatter was of mt-DNA haplogroup W. Her closest mt-DNA match (perfect match to Richard's HVR1 and HVR2) is with an individual whose earliest known maternal ancestor was a woman named Karen Knudsdatter. Karen was married January 13, 1734 in Nord-Aurdal, Oppland, Sweden, to Enblebret Tostensen. Statistically speaking, Ingeborg Pedersdatter and Karen Knudsdatter are likely to share a common direct maternal ancestor, perhaps 60 generations back, somewhere in Europe. However, the close proximity between Eigersund and Nord-Aurdal (approximately 300 miles as the crow flies) suggests that the most recent common mt-DNA mutation probably occurred much closer to the present, and specifically in a woman who resided in Norway.
My Direct Maternal Line
Pictured is my great-grandmother Eva Rice. Eva is my earliest known direct maternal ancestor. She was born around October 31, 1883 in Lithuania. Her name at birth was Ieva Daukšaitė, shortened to Eva Douse after immigrating to the United States. She took the surname Rice after marrying my great-grandfather Peter Rice, a descendant of the Lithuanian Račius family. Eva's father was Jonas Daukša. Her mother, whose name is not currently known, died in Lithuania, probably giving birth to Eva. For a full biography of Eva Rice, see my previous article on Familypedia.
Based on a sample of my mitochondrial DNA, Eva was of mt-DNA haplogroup T2. My closest mt-DNA match (perfect match to my HVR1 and HVR2) is with an individual from Bulgaria, whose earliest known maternal ancestor had the Slavic maiden name Fiyodorova. My next two closest matches (perfect match to my HVR1, but they did not test their HVR2) are with individuals whose earliest known maternal ancestors are of Polish origin, with maiden names Borek and Stefineski. The common maternal ancestor of all four lineages was likely a woman who lived in Poland or the Ukrane less than 2000 years ago. Although Lithuanians speak a Baltic language, it is not surprising that a person from Lithuania would have Slavic, and particularly Polish ancestry. Based on y-chromosomal DNA studies of Baltic populations, 38% of Lithuanians are of y-DNA haplogroup R1a, as are 55% of the population of Poland. (Another 42% of Lithuanians are of y-DNA haplogroup N, indicating relationship to Uralic speakers.)
Mt-DNA haplogroup T (ancestral to T2) likely arrived in Poland long before Indo-European y-DNA haplogroup R1a, as the incidence of haplogroup T corresponds more directly with R1 in Europe, generally, than it does with either of its R1a or earlier arriving R1b subclades individually (which, for reasons beyond the scope of this article, does not imply that mt-DNA haplogroup T and y-DNA haplogroup R1 are necessarily closely related).
The individual woman who first had the mutation that defined mt-DNA haplogroup T, my direct maternal ancestor, has been given the nickname "Tara" by geneticists. Tara probably lived in the Near East, more than 45,000 years ago. Tara was also a direct maternal ancestor of outlaw Jesse James, and Tsar Nicholas II. The International Society of Genetic Genealogy places haplogroup T2 (shared by Jesse James, Tsar Nicholas II and myself) in Europe sometime between the late Stone Age and 8000 BC.
My Father's Direct Maternal Line
This is a photograph of my great-grandmother Maria Freudenberg (1895-1987), the common direct maternal ancestor of my father and my cousin Richard Norton who shared his mt-DNA results with me. Maria was born Maria Elizabeth Winblad. Maria's mother was Salmine Sofie Severine Olsdatter (1862-1914), who was born in Farsund, Vest-Agder, Norway. Salmine's mother was Thea Johanne Tostensdatter (1825-1865), born in Kristiansand, Vest-Agder, Norway. Thea's mother was Johanne Jacobsdatter (1795-?), born in Eigersund, Rogaland, Norway. Johanne's mother was Gitlaug Tollaksdatter (1773-1806), born in Hå, Rogaland, Norway. Gitlaug's mother was Ingeborg Pedersdatter (c1747-?), believed to have been born in Eigersund.
Based on a sample of my cousin's mitochondrial DNA, Ingeborg Pedersdatter was of mt-DNA haplogroup W. Her closest mt-DNA match (perfect match to Richard's HVR1 and HVR2) is with an individual whose earliest known maternal ancestor was a woman named Karen Knudsdatter. Karen was married January 13, 1734 in Nord-Aurdal, Oppland, Sweden, to Enblebret Tostensen. Statistically speaking, Ingeborg Pedersdatter and Karen Knudsdatter are likely to share a common direct maternal ancestor, perhaps 60 generations back, somewhere in Europe. However, the close proximity between Eigersund and Nord-Aurdal (approximately 300 miles as the crow flies) suggests that the most recent common mt-DNA mutation probably occurred much closer to the present, and specifically in a woman who resided in Norway.
Saturday, November 5, 2011
My Ancestors From Farsund, Norway

Generation 1: Salmine Sofie Severine Olsdatter immigrated 1884 to New York City from Farsund, Vest-Agder, Norway, shortly after her marriage to John Edward Winblad in 1882. John Winblad was a native of Sweden. Salmine is believed to have been born on the Brækne farm, near Vanse, Farsund. Her baptismal record indicates a birth date of March 27, 1862, and parents Ole Mattias Pedersen and Tea Johanne Torstensdatter. Salmine was baptized in Lista (a former municipality which includes Vanse), June 8, 1862. By 1900, Salmine resided in Jersey City, New Jersey. She eventually returned to Norway, where she died December 18, 1914.
Generation 2: Ole Mathias Pedersen was born November 14, 1822 on the Log farm near Herad, Farsund. He was baptized November 17 in Herad. His baptismal record indicates parents Peder Andreas Hansen and Maren Sophia Olsdatter. Ole married Thea Johanne Tostensdatter from nearby Vetteland, Kristiansand, Vest-Agder, on October 8, 1848. The marriage record confirms the name of Ole's father, i.e. Peder Andreas Hansen. Ole died August 24, 1914 in Klungeland, Vanse. By occupation, Ole was a baker.
Generation 3: Peder Andreas Hansen, also of Log, was baptized January 3, 1790 in Herad. His baptismal record only lists his father, Hans Hansen, although further research has shown that his mother was Pernille Helene Pedersdatter of Meberg, Kvinesdal, Vest-Agder. Peder married Maren Sophie Olsdatter of Gullestad, Kvinesdal, December 28, 1811 in Herad.
Generation 4: Hans Hansen, also of Log, was born December 4, 1749, and baptized in Herad, December 5, 1749. Hans' parents were Hans Hansen and Ingri Ingvorsdatter, both of Farsund.
Generation 5: Baptismal records for both Hans Hansen and Ingri Ingvorsdatter have been located, indicating that Hans Hansen was baptized May 15, 1729 in Herad, son of Hans Larsen, and that Ingri Ingvorsdatter was baptized November 25, 1726 in Herad, daughter of Ingvor Rassmusen. At this point, little is known about Hans Larsen, Ingvor Rassmusen, or their wives. However, searching through baptismal records in Herad has revealed the names of additional children. In addition to Hans Hansen (1729), Hans Larsen was also father to Peder Hansen (1727), Johannes Hansen (1731) and Christen Hansen (1734). In addition to Ingri Ingvorsdatter (1726), Ingvor Rasmusen was also father to Rasmus Ingvorsen (1730) and Engel Ingvorsdatter (1734).
Generation 6: Ingvor Rasmusen had two known siblings, both listed as children of "Rasmus of Log" in their baptismal records, Guri Rasmusdatter (1696-1697) and Hans Rasmussen (1698-1699). Neither of these siblings survived to maturity.
Monday, October 17, 2011
Flood Barriers Hold Firm in Shielding Bangkok - ABC News
According to this news article from ABC, the residents of Bangkok are hoping that a series of dams, canals and tunnels spares the city from this year's intense flooding that has caused catastrophic damage to Thailand's rice-basket region, just upstream from Bangkok. Recent news reports have shown footage of residents in Ayutthaya using boats to travel down the city's streets.

This seems like deja vu. Exactly four years ago, I wrote this blog post: Flooding in Phitsanulok, Thailand. My wife says that her family in Ban Dong Phayom (Phitsanulok Province) has been hit hard by the flooding. Houses are raised on piles in the rural farming village where she grew up. Even so, her brothers and sisters have reported that the water level has risen enough to enter a large percentage of the elevated dwellings.

Typical home in Ban Dong Phayom. Photo by Kevin Borland.
My newphew's wife just sent me these photos she took near her home in Nonthaburi. Nonthaburi is just northwest of Bangkok.








This seems like deja vu. Exactly four years ago, I wrote this blog post: Flooding in Phitsanulok, Thailand. My wife says that her family in Ban Dong Phayom (Phitsanulok Province) has been hit hard by the flooding. Houses are raised on piles in the rural farming village where she grew up. Even so, her brothers and sisters have reported that the water level has risen enough to enter a large percentage of the elevated dwellings.
Typical home in Ban Dong Phayom. Photo by Kevin Borland.
My newphew's wife just sent me these photos she took near her home in Nonthaburi. Nonthaburi is just northwest of Bangkok.
Saturday, October 15, 2011
My Direct Paternal Ancestry
Growing up, I knew very little about my direct paternal ancestry. I first met my paternal great grandfather Weldon Earl Borland after the death of my grandfather John Earl Borland in 1986. After college, I began working on creating a family tree, beginning with information I obtained from interviewing Weldon. I have since continued compiling information regarding the history of the Borland family, and recently had my Y chromosomal DNA tested. The following is the history of my paternal ancestral line, generation by generation:
Generation 1: Kevin Borland
I was born January 20, 1975, in Jersey City, New Jersey, to parents Steven Thomas and Kathleen (Szczesny) Borland. Shortly after my birth, my family moved to Morris County, New Jersey, where we lived for three years before relocating further northwest to Hardyston, Sussex County.


My childhood homes in Lake Hiawatha, Morris County (left) and Hardyston, Sussex County (right)
I currently reside in Arlington, Virginia, with my wife Thidawan and my stepson Jaras. I am an attorney by profession.

Kevin Borland family
Generation 2: Steven Thomas Borland
My father was born August 7, 1950, also in Jersey City, to parents John Earl and Helen Eloise (Freudenberg) Borland. He grew up in Jersey City, and remained in New Jersey until retiring to Florida about 10 years ago. Steven served in the United States Army, and was deployed to Germany during the Vietnam War. Upon his return to New Jersey, he worked for the United Parcel Service (UPS) most of his life, until he began suffering from leukemia. He died in Tampa June 15, 2010, from complications arising from a bone marrow transplant, after which he was buried in Leesville, Ohio, a few rows away from his father's grave.


Steven Thomas Borland's high school yearbook photo (left) and pictured with his two sons Kevin and Steven in Florida (right)
Generation 3: John Earl Borland
John Earl Borland was born May 16, 1924, in the now-defunct village of New Hagerstown, Carroll County, Ohio, to parents Weldon Earl and Elizabeth Marie (Forbes) Borland. John's parents divorced prior to his birth, and John was raised by his mother Elizabeth on her parents' farm in New Hagerstown. John served in the United States Navy during World War II, after which he relocated to Jersey City, where he met his first wife, my grandmother Helen. John worked most of his adult life at the Ballentine Brewery in Newark, New Jersey. John and Helen eventually divorced, and John remarried to Geraldine (Winblad) Van Deusen. Upon John's retirement, he returned to Ohio with Geraldine, where he died of lung cancer, November 10, 1986.


John Borland's high school graduation photo (left) and pictured with wife Helen, stepson Michael and sons John and Steven (right)
Generations 4: Weldon Earl Borland
Weldon Earl Borland was born July 3, 1906 in Bowerston, Harrison County, Ohio, to parents James Couthren and Lizzie Alberta (Miller) Borland. Weldon grew up in Bowerston, but as an adult relocated to Akron, Ohio, where he worked for Goodyear. After some time, he left Goodyear and started his own business called "Volume Sewing." Weldon and his employees manufactured a variety of sewn items ranging from army uniforms to boat covers. Weldon married briefly my great-grandmother Elizabeth Marie Forbes, and later Vivian Kniseley. My great-uncle Jeffrey James Borland is a son from Weldon's second marriage. Weldon died March 2, 2002, while he was hospitalized for treatment of pneumonia, and was buried in Long View Cemetery in Bowerston, as are both of his parents. He was 95 years old at the time of his death.


Weldon Borland in his youth (left) and later in life pictured with his long-time companion Rose Simmons (right)
Generation 5: James Crouthen Borland
James Crouthen Borland was born September 25, 1877, also in Bowerston, to parents James II and Catherine Jane (Walker) Borland. He was the owner of "J.C. Borland Wholesale and Retail," a general store in Bowerston. James and wife Lizzie Alberta (Miller) Borland had four children, two of which survived to maturity, my great-grandfather Weldon, and his sister Ruth Eleanor Borland. James died April 19, 1943 in Bowerston, of a cerebral hemorrhage secondary to chronic alcoholism. At the time of James' death, he and Lizzie were divorced.


James Borland and family behind the counter at J.C. Borland Wholesale and Retail (left), and James' letterhead (right)
Generation 6: James Borland II
James Borland II was born October 2, 1835 in Orange Township, Carroll County, Ohio, probably in or near New Hagerstown. His parents were James and Mary (McQuiston) Borland. He married Catherine Jane Walker and they had two children, Charles Oliver Borland (1871-1931) and James Crouthen Borland (1877-1943). James was a farmer by occupation, and he served as a volunteer soldier in the United States Civil War. His Civil War rifle remains in the family. James died May 11, 1903 in Bowerston.


James Borland II with his horse (left), grave of James and Catherine Borland at Long View Cemetery (right)
Generation 7: James Borland
James Borland was born 1792 in or near Westmoreland County, Pennsylvania, to parents Samuel and Lydia (Gregg) Borland. He married Mary McQuiston of Butler, Pennsylvania, with whom he fathered at least six children. A McQuiston family history states that James was a music teacher in his youth. Census data reveals that he was a farmer, later in life. James lived until at least 1850, although his grave has not been located. It is suspected that he may be buried in New Hagerstown.
Generation 8: Samuel Borland
Samuel Borland was born 1748 in County Antrim, Northern Ireland, to parents John and Rachel Borland. After immigrating to the United States prior to 1783, he married Lydia Gregg in Pennsylvania and had 11 known children. Samuel and his family resided in Manor of Denmark, near Export, Westmoreland County, Pennsylvania, since at least 1800, where Samuel built the first stone house in the Manor Valley. Samuel died 1811, after which Lydia obtained a land patent in Carroll County, Ohio, where subsequent Borland generations settled. Samuel and Lydia are buried in Congruity, Salem Township, Westmoreland County. Lydia's suspected brother David Gregg was a great-grandfather of United States President Harry S Truman.

Samuel Borland residence in Manor of Denmark (left), Samuel and Lydia Borland graves (right)
Generation 9: John Borland
Based on the year of his first marriage (1738), John Borland must have been born prior to 1720. Based on the names of his children, his father was likely named James. John was the progenitor of the Borland family of the Kilraughts Parish in County Antrim. He first married a woman by the name of Ann in Lisburn, by whom he fathered four children. After Ann's death, prior to 1745, he remarried to a woman named Rachel, widow of a Mr. Moore, by whom he fathered an additional 9 children. While some of the children immigrated to the United States, others remained in and around Kilraughts. John was a farmer by occupation. He died around 1778 in or near Kilraughts. The location of John's birth is presently unknown, although one might speculate that since he first married in Lisburn, he was probably born somewhere in the vicinity of Belfast.

Borland residence in Kilraughts
Generation 10: James Borland
The name James is based on the fact that John Borland (of generation 9) named his first-born son James. The custom at the time, both in Scotland and Northern Ireland, was to name one's first-born son after the child's paternal grandfather. James would have been born circa 1685, probably in or near Belfast. In addition to his son John of Kilraughts, he also fatherd a son Andrew who resided in Ballymoney. My direct line of paternal ancestry can only be traced as far as this 10th generation using conventional genealogical methods, i.e. through the discovery and review of historic documents, gravestones, published family histories, etc.
Generations 11-12: Borlands of Belfast
The similarity between my Y-DNA and the Y-DNA of David Hunter Borland of Pittsburgh, Pennsylvania, suggests that the Borlands of Clondavaddog, County Donegal, descend from a very close relative of James Borland (of generation 10). Furthermore, based on the small number of Borland families in Ulster (the northernmost of Ireland's 4 provinces) enumerated in the 1796 list of Irish flax growers, clustered almost entirely near Clondavaddog and Ballymoney, it would appear that either James' father or grandfather was likely the immigrant ancestor of all of the early Borland families of Ulster, having come to Belfast from Scotland (Borland being a Scottish surname). According to the Wikipedia Article entitled "History of Belfast," after the Irish Rebellion of 1641, many Scots who had come to Ulster as part of the Scottish army sent to put down the rebellion, settled in Belfast after the Irish Confederate Wars." A Borland ancestor of about the 12th generation would have been just the right age (having likely been born around the 1620s) to have been one of the Scottish soldiers.
Generations 13-15: Borlands of Strathclyde
According to Ancestry.com, my closest Y-DNA match (using 46 short tandem repeat markers) is to a Randy Scott Borland, who, like me, resides in Northern Virginia. Ancestry.com estimates that we are probably related within 15 generations. Although I have been thusfar unable to contact Randy, I independently constructed his family tree. Upon doing so, I learned that he descends from a line of Borlands that straddles the parishes of Galston and Loudoun, in the Strathclyde region of Scotland. His Borland line does not pass through Ireland. This suggests that that our distinct branches of the Borland family both descend from a common Borland ancestor who lived in the Strathclyde region of Scotland, possibly born around the 1530s (based on an average generation gap of 30 years). Unfortunately, the church records of Galston and Loudoun only go back to the 1680s, where in Loudoun, for example, in 1684 a child named Anna was baptized, with the name Archiblad Borland listed as her father.
Generations 16-18: Origin of the Borland Surname
In Ancestry.com's ranking of genetic separation of Y-DNA donors, once we reach STR matches estimated within approximately 19 generations, all matches have surnames other than Borland. This suggests that the Borland surname was adopted in my direct paternal line between generations 16 and 18. Again using an estimated average generation gap of 30 years, that brings us to the time period between the 1440s and 1500s. My first ancestor to have used the name Borland would have spoken an Anglic language, probably either Scots or Middle English (ancestral to the modern English language).
Generations 19-21: Beyond Borland and Beyond Scotland
STR matches estimated to converge with my paternal lineage at generations 19 and 21 carry the surnames Shaw and Fielder, respectively. Both of these surnames, like the name Borland, are of Anglic origin, suggesting that the Borlands, Shaws and Fielders, shared a common Anglic-speaking direct paternal ancestor born around 1350, who likely lived in England (since Shaw and Borland are Scottish names, whereas Fielder, a line which split off from my lineage earlier is of English origin). Interestingly, all three names are also topographic surnames. It would appear that the names Borland, Shaw and Fielder were adopted by three branches of the same family, prior to which (before generation 18) the family may not have employed the use of a surname. The migration from England to Scotland likely occurred around generation 20.
Generation 22: Brief Passing Through Cornwall
A 12/12 STR match, found on ySearch.org, with an individual bearing the surname Pennock suggests a 90% chance of a direct paternal relation within 23 generations. Pennock is a Cornish name, indicating that the Borland paternal ancestor born around 1320 may have lived in Cornwall, in southwestern England. Pennock, once again, is a topographic surname, deriving its name from Pignocshire (pronounced with a silent "g") in Cornwall.
Generation 23: From Gascony to England During the Hundred Years' War
Another 12/12 STR match from ySearch.org reveals a link within 23 generations to an individual with the surname "de Ayala" who has traced his paternal line back to Gascony, a historically Basque region in southwestern France. "De Ayala" is another topographic surname originating in Ayala, a village in Basque Country, Spain. This suggests that my 20x great-grandfather, born around the year 1290, lived among the Basque, in either northwestern Spain or southwestern France. I suspect he resided in Gascony, since that would explain the migration in the next generation or so to Cornwall. This has to do with the Edwardian War being fought in Gascony from 1337-1360 (the first phase of the Hundred Years' War). At that time, Gascony was English territory. It would not be surprising if some citizens of Gascony may have moved to other, perhaps more peaceful, English territories during the war. Cornwall is the closest portion of present-day England to Gascony, geographically, perhaps making it an obvious choice for my ancestor, even if the trip was almost 700 miles by sea. If this theory is correct, than the name "de Ayala" may have been the surname used by my direct paternal ancestors prior to the adoption of an Anglic language. This would explain the various Anglic topographic surnames used by different branches of the family in later generations. Perhaps each branch of the family switched from "de Ayala" to a new Anglic topographic surname upon adoption of an Anglic language. This would also explain why the surname Borland seems to have arisen at a time long after surnames were being commonly used in Scotland.
Generations 24-28: Basque Country
A more distant STR match, estimated by Ancestry.com to converge with my paternal lineage at generation 28 (mid 12th century), further supports a Basque Country homeland of the paternal ancestors of the Borlands. At this level, the surname Zurita is added to the list of surnames sharing a common paternal origin. Zurita is an Aragonese surname, originating in northern Spain. Y-DNA STR analysis appears to indicate that my direct paternal line resided in the Pyrenees by the 13th century, where Navarro-Aragonese languages were spoken, in addition to Basque. Muslims would have been more likely to have spoken the Navarro-Aragonese languages, since they did not descend from the indigenous Basque people.
Generation 29: Salamanca to Basque Country
A 31/37 STR match in the Sorenson Molecular Genealogy Foundation database with an individual with the topographic surname Bejar suggests my paternal ancestral line may have resided in or near Salamanca in the 29th generation. Béjar is the name of a town in Salamanca. The migration from Salamanca to the Basque Country in the Pyrenees was likely a result of the Reconquista, a gradual process in the Middle Ages during which several Christian kingdoms succeeded in retaking the Muslim-controlled areas of the Iberian Peninsula broadly known as Al-Andalus. The paternal ancestors of the de Ayala/Borland family , were likely Muslims (Moors) who retreated to the mountains in northwestern Spain as a result of the Reconquista. (See below for evidence supporting this theory.) According to Wikipedia, "The main phase of the Reconquista was completed by 1249, after the Battle of Las Navas de Tolosa, when the sole remaining Muslim state in Iberia, the Emirate of Granada, became a vassal state of the Christian Crown of Castile." This fits very well into our generational timeline. The animated map below, provided by the Norwegian historian Tolke, depicts the timeline of the Reconquista. The retracting green portion of the animation represents the portion of the Iberian Peninsula dominated by the Moors.

Generations 30-34: One Hundred Fifty More Years in Spain
Continuing on through generation 34 (early 10th century), the surnames Salinero, Levario, Rodriguez and Bermudez appear among my STR matches, indicating the presence of my paternal line in Spain for over two centuries. Based on these patronymic and occupational surnames, however, a precise region in Spain cannot be identified.
Generations 35-40: Two More Centuries in Spain
Beyond 35 generations, STR comparison ceases to be a particularly useful method of tracing paternal migration, and we must turn to single-nucleotide polymorphism. While I have not personally taken a deep SNP y-dna haplogroup test, fortunately, my approximately 30th paternal cousin Armondo C. Rodriguez has. His results reveal that he, as well as the Borlands, are of y-DNA haplogroup E1b1b1b1b (M183), indicating recent North African ancestry. We then turn to the history of Spain, and discover that in the early 8th century (around generation 40 on our timeline), nearly all of the Iberian Peninsula was conquered (711–718) by largely Moorish Muslim armies from North Africa. The Moors referred to the conquered peninsula as Al-Andalus. My paternal 37x great-grandfather would have spoken Andalusian Arabic.
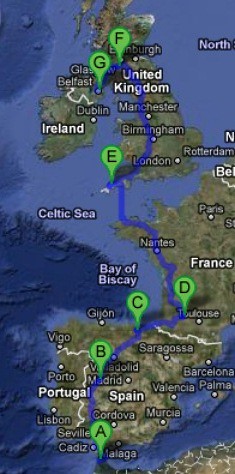
Map showing migration route from Northern Africa to Northern Ireland, as discussed thusfar, with nodes at A: Morocco, B: Salamanca, C: Basque Country, D: Gascony, E: Cornwall, F: Strathclyde and G: Ulster.
Generations 41-185: Over Four Thousand Years in the Maghreb
Y-DNA haplogroup E1b1b1b1b (M183) is believed to have emerged in the Maghreb, or northwestern Africa, around 5,600 years ago, which would suggest that my paternal ancestors spent over 4,000 years living in the Sahara. M183 is sometimes referred to as the Berber marker, since in some groups of Berbers, nearly 100% of the population exhibit this mutation. The Moors who invaded Spain were largely of Berber ethnicity.
Generations 186-750: From the Nile to the Sahara
Haplogroup E1b1b (M215), ancestral to E1b1b1b1b (M183) is believed to have emerged in eastern Africa, around 22,400 years ago, perhaps along the Nile in present-day Ethiopia or Sudan. 750 generations ago, my direct paternal ancestor may have spoken an ancient language, ancestral to the modern Nilo-Saharan languages.
Generations 751-1200: From the Ethiopian Highlands
Haplogroup E1b1 (P2/PN2), ancestral to E1b1b (M215) is believed to have emerged in the Ethiopian highlands, around 35,000 years ago. 1200 generations ago, my direct paternal ancestor may have spoken a language not only ancestral to the Nilo-Saharan languages, but also to the Niger-Congo and Mande languages.
Generation 1: Kevin Borland
I was born January 20, 1975, in Jersey City, New Jersey, to parents Steven Thomas and Kathleen (Szczesny) Borland. Shortly after my birth, my family moved to Morris County, New Jersey, where we lived for three years before relocating further northwest to Hardyston, Sussex County.
My childhood homes in Lake Hiawatha, Morris County (left) and Hardyston, Sussex County (right)
I currently reside in Arlington, Virginia, with my wife Thidawan and my stepson Jaras. I am an attorney by profession.
Kevin Borland family
Generation 2: Steven Thomas Borland
My father was born August 7, 1950, also in Jersey City, to parents John Earl and Helen Eloise (Freudenberg) Borland. He grew up in Jersey City, and remained in New Jersey until retiring to Florida about 10 years ago. Steven served in the United States Army, and was deployed to Germany during the Vietnam War. Upon his return to New Jersey, he worked for the United Parcel Service (UPS) most of his life, until he began suffering from leukemia. He died in Tampa June 15, 2010, from complications arising from a bone marrow transplant, after which he was buried in Leesville, Ohio, a few rows away from his father's grave.
Steven Thomas Borland's high school yearbook photo (left) and pictured with his two sons Kevin and Steven in Florida (right)
Generation 3: John Earl Borland
John Earl Borland was born May 16, 1924, in the now-defunct village of New Hagerstown, Carroll County, Ohio, to parents Weldon Earl and Elizabeth Marie (Forbes) Borland. John's parents divorced prior to his birth, and John was raised by his mother Elizabeth on her parents' farm in New Hagerstown. John served in the United States Navy during World War II, after which he relocated to Jersey City, where he met his first wife, my grandmother Helen. John worked most of his adult life at the Ballentine Brewery in Newark, New Jersey. John and Helen eventually divorced, and John remarried to Geraldine (Winblad) Van Deusen. Upon John's retirement, he returned to Ohio with Geraldine, where he died of lung cancer, November 10, 1986.
John Borland's high school graduation photo (left) and pictured with wife Helen, stepson Michael and sons John and Steven (right)
Generations 4: Weldon Earl Borland
Weldon Earl Borland was born July 3, 1906 in Bowerston, Harrison County, Ohio, to parents James Couthren and Lizzie Alberta (Miller) Borland. Weldon grew up in Bowerston, but as an adult relocated to Akron, Ohio, where he worked for Goodyear. After some time, he left Goodyear and started his own business called "Volume Sewing." Weldon and his employees manufactured a variety of sewn items ranging from army uniforms to boat covers. Weldon married briefly my great-grandmother Elizabeth Marie Forbes, and later Vivian Kniseley. My great-uncle Jeffrey James Borland is a son from Weldon's second marriage. Weldon died March 2, 2002, while he was hospitalized for treatment of pneumonia, and was buried in Long View Cemetery in Bowerston, as are both of his parents. He was 95 years old at the time of his death.
Weldon Borland in his youth (left) and later in life pictured with his long-time companion Rose Simmons (right)
Generation 5: James Crouthen Borland
James Crouthen Borland was born September 25, 1877, also in Bowerston, to parents James II and Catherine Jane (Walker) Borland. He was the owner of "J.C. Borland Wholesale and Retail," a general store in Bowerston. James and wife Lizzie Alberta (Miller) Borland had four children, two of which survived to maturity, my great-grandfather Weldon, and his sister Ruth Eleanor Borland. James died April 19, 1943 in Bowerston, of a cerebral hemorrhage secondary to chronic alcoholism. At the time of James' death, he and Lizzie were divorced.
James Borland and family behind the counter at J.C. Borland Wholesale and Retail (left), and James' letterhead (right)
Generation 6: James Borland II
James Borland II was born October 2, 1835 in Orange Township, Carroll County, Ohio, probably in or near New Hagerstown. His parents were James and Mary (McQuiston) Borland. He married Catherine Jane Walker and they had two children, Charles Oliver Borland (1871-1931) and James Crouthen Borland (1877-1943). James was a farmer by occupation, and he served as a volunteer soldier in the United States Civil War. His Civil War rifle remains in the family. James died May 11, 1903 in Bowerston.
James Borland II with his horse (left), grave of James and Catherine Borland at Long View Cemetery (right)
Generation 7: James Borland
James Borland was born 1792 in or near Westmoreland County, Pennsylvania, to parents Samuel and Lydia (Gregg) Borland. He married Mary McQuiston of Butler, Pennsylvania, with whom he fathered at least six children. A McQuiston family history states that James was a music teacher in his youth. Census data reveals that he was a farmer, later in life. James lived until at least 1850, although his grave has not been located. It is suspected that he may be buried in New Hagerstown.
Generation 8: Samuel Borland
Samuel Borland was born 1748 in County Antrim, Northern Ireland, to parents John and Rachel Borland. After immigrating to the United States prior to 1783, he married Lydia Gregg in Pennsylvania and had 11 known children. Samuel and his family resided in Manor of Denmark, near Export, Westmoreland County, Pennsylvania, since at least 1800, where Samuel built the first stone house in the Manor Valley. Samuel died 1811, after which Lydia obtained a land patent in Carroll County, Ohio, where subsequent Borland generations settled. Samuel and Lydia are buried in Congruity, Salem Township, Westmoreland County. Lydia's suspected brother David Gregg was a great-grandfather of United States President Harry S Truman.
Samuel Borland residence in Manor of Denmark (left), Samuel and Lydia Borland graves (right)
Generation 9: John Borland
Based on the year of his first marriage (1738), John Borland must have been born prior to 1720. Based on the names of his children, his father was likely named James. John was the progenitor of the Borland family of the Kilraughts Parish in County Antrim. He first married a woman by the name of Ann in Lisburn, by whom he fathered four children. After Ann's death, prior to 1745, he remarried to a woman named Rachel, widow of a Mr. Moore, by whom he fathered an additional 9 children. While some of the children immigrated to the United States, others remained in and around Kilraughts. John was a farmer by occupation. He died around 1778 in or near Kilraughts. The location of John's birth is presently unknown, although one might speculate that since he first married in Lisburn, he was probably born somewhere in the vicinity of Belfast.
Borland residence in Kilraughts
Generation 10: James Borland
The name James is based on the fact that John Borland (of generation 9) named his first-born son James. The custom at the time, both in Scotland and Northern Ireland, was to name one's first-born son after the child's paternal grandfather. James would have been born circa 1685, probably in or near Belfast. In addition to his son John of Kilraughts, he also fatherd a son Andrew who resided in Ballymoney. My direct line of paternal ancestry can only be traced as far as this 10th generation using conventional genealogical methods, i.e. through the discovery and review of historic documents, gravestones, published family histories, etc.
Generations 11-12: Borlands of Belfast
The similarity between my Y-DNA and the Y-DNA of David Hunter Borland of Pittsburgh, Pennsylvania, suggests that the Borlands of Clondavaddog, County Donegal, descend from a very close relative of James Borland (of generation 10). Furthermore, based on the small number of Borland families in Ulster (the northernmost of Ireland's 4 provinces) enumerated in the 1796 list of Irish flax growers, clustered almost entirely near Clondavaddog and Ballymoney, it would appear that either James' father or grandfather was likely the immigrant ancestor of all of the early Borland families of Ulster, having come to Belfast from Scotland (Borland being a Scottish surname). According to the Wikipedia Article entitled "History of Belfast," after the Irish Rebellion of 1641, many Scots who had come to Ulster as part of the Scottish army sent to put down the rebellion, settled in Belfast after the Irish Confederate Wars." A Borland ancestor of about the 12th generation would have been just the right age (having likely been born around the 1620s) to have been one of the Scottish soldiers.
Generations 13-15: Borlands of Strathclyde
According to Ancestry.com, my closest Y-DNA match (using 46 short tandem repeat markers) is to a Randy Scott Borland, who, like me, resides in Northern Virginia. Ancestry.com estimates that we are probably related within 15 generations. Although I have been thusfar unable to contact Randy, I independently constructed his family tree. Upon doing so, I learned that he descends from a line of Borlands that straddles the parishes of Galston and Loudoun, in the Strathclyde region of Scotland. His Borland line does not pass through Ireland. This suggests that that our distinct branches of the Borland family both descend from a common Borland ancestor who lived in the Strathclyde region of Scotland, possibly born around the 1530s (based on an average generation gap of 30 years). Unfortunately, the church records of Galston and Loudoun only go back to the 1680s, where in Loudoun, for example, in 1684 a child named Anna was baptized, with the name Archiblad Borland listed as her father.
Generations 16-18: Origin of the Borland Surname
In Ancestry.com's ranking of genetic separation of Y-DNA donors, once we reach STR matches estimated within approximately 19 generations, all matches have surnames other than Borland. This suggests that the Borland surname was adopted in my direct paternal line between generations 16 and 18. Again using an estimated average generation gap of 30 years, that brings us to the time period between the 1440s and 1500s. My first ancestor to have used the name Borland would have spoken an Anglic language, probably either Scots or Middle English (ancestral to the modern English language).
Generations 19-21: Beyond Borland and Beyond Scotland
STR matches estimated to converge with my paternal lineage at generations 19 and 21 carry the surnames Shaw and Fielder, respectively. Both of these surnames, like the name Borland, are of Anglic origin, suggesting that the Borlands, Shaws and Fielders, shared a common Anglic-speaking direct paternal ancestor born around 1350, who likely lived in England (since Shaw and Borland are Scottish names, whereas Fielder, a line which split off from my lineage earlier is of English origin). Interestingly, all three names are also topographic surnames. It would appear that the names Borland, Shaw and Fielder were adopted by three branches of the same family, prior to which (before generation 18) the family may not have employed the use of a surname. The migration from England to Scotland likely occurred around generation 20.
Generation 22: Brief Passing Through Cornwall
A 12/12 STR match, found on ySearch.org, with an individual bearing the surname Pennock suggests a 90% chance of a direct paternal relation within 23 generations. Pennock is a Cornish name, indicating that the Borland paternal ancestor born around 1320 may have lived in Cornwall, in southwestern England. Pennock, once again, is a topographic surname, deriving its name from Pignocshire (pronounced with a silent "g") in Cornwall.
Generation 23: From Gascony to England During the Hundred Years' War
Another 12/12 STR match from ySearch.org reveals a link within 23 generations to an individual with the surname "de Ayala" who has traced his paternal line back to Gascony, a historically Basque region in southwestern France. "De Ayala" is another topographic surname originating in Ayala, a village in Basque Country, Spain. This suggests that my 20x great-grandfather, born around the year 1290, lived among the Basque, in either northwestern Spain or southwestern France. I suspect he resided in Gascony, since that would explain the migration in the next generation or so to Cornwall. This has to do with the Edwardian War being fought in Gascony from 1337-1360 (the first phase of the Hundred Years' War). At that time, Gascony was English territory. It would not be surprising if some citizens of Gascony may have moved to other, perhaps more peaceful, English territories during the war. Cornwall is the closest portion of present-day England to Gascony, geographically, perhaps making it an obvious choice for my ancestor, even if the trip was almost 700 miles by sea. If this theory is correct, than the name "de Ayala" may have been the surname used by my direct paternal ancestors prior to the adoption of an Anglic language. This would explain the various Anglic topographic surnames used by different branches of the family in later generations. Perhaps each branch of the family switched from "de Ayala" to a new Anglic topographic surname upon adoption of an Anglic language. This would also explain why the surname Borland seems to have arisen at a time long after surnames were being commonly used in Scotland.
Generations 24-28: Basque Country
A more distant STR match, estimated by Ancestry.com to converge with my paternal lineage at generation 28 (mid 12th century), further supports a Basque Country homeland of the paternal ancestors of the Borlands. At this level, the surname Zurita is added to the list of surnames sharing a common paternal origin. Zurita is an Aragonese surname, originating in northern Spain. Y-DNA STR analysis appears to indicate that my direct paternal line resided in the Pyrenees by the 13th century, where Navarro-Aragonese languages were spoken, in addition to Basque. Muslims would have been more likely to have spoken the Navarro-Aragonese languages, since they did not descend from the indigenous Basque people.
Generation 29: Salamanca to Basque Country
A 31/37 STR match in the Sorenson Molecular Genealogy Foundation database with an individual with the topographic surname Bejar suggests my paternal ancestral line may have resided in or near Salamanca in the 29th generation. Béjar is the name of a town in Salamanca. The migration from Salamanca to the Basque Country in the Pyrenees was likely a result of the Reconquista, a gradual process in the Middle Ages during which several Christian kingdoms succeeded in retaking the Muslim-controlled areas of the Iberian Peninsula broadly known as Al-Andalus. The paternal ancestors of the de Ayala/Borland family , were likely Muslims (Moors) who retreated to the mountains in northwestern Spain as a result of the Reconquista. (See below for evidence supporting this theory.) According to Wikipedia, "The main phase of the Reconquista was completed by 1249, after the Battle of Las Navas de Tolosa, when the sole remaining Muslim state in Iberia, the Emirate of Granada, became a vassal state of the Christian Crown of Castile." This fits very well into our generational timeline. The animated map below, provided by the Norwegian historian Tolke, depicts the timeline of the Reconquista. The retracting green portion of the animation represents the portion of the Iberian Peninsula dominated by the Moors.
Generations 30-34: One Hundred Fifty More Years in Spain
Continuing on through generation 34 (early 10th century), the surnames Salinero, Levario, Rodriguez and Bermudez appear among my STR matches, indicating the presence of my paternal line in Spain for over two centuries. Based on these patronymic and occupational surnames, however, a precise region in Spain cannot be identified.
Generations 35-40: Two More Centuries in Spain
Beyond 35 generations, STR comparison ceases to be a particularly useful method of tracing paternal migration, and we must turn to single-nucleotide polymorphism. While I have not personally taken a deep SNP y-dna haplogroup test, fortunately, my approximately 30th paternal cousin Armondo C. Rodriguez has. His results reveal that he, as well as the Borlands, are of y-DNA haplogroup E1b1b1b1b (M183), indicating recent North African ancestry. We then turn to the history of Spain, and discover that in the early 8th century (around generation 40 on our timeline), nearly all of the Iberian Peninsula was conquered (711–718) by largely Moorish Muslim armies from North Africa. The Moors referred to the conquered peninsula as Al-Andalus. My paternal 37x great-grandfather would have spoken Andalusian Arabic.
Map showing migration route from Northern Africa to Northern Ireland, as discussed thusfar, with nodes at A: Morocco, B: Salamanca, C: Basque Country, D: Gascony, E: Cornwall, F: Strathclyde and G: Ulster.
Generations 41-185: Over Four Thousand Years in the Maghreb
Y-DNA haplogroup E1b1b1b1b (M183) is believed to have emerged in the Maghreb, or northwestern Africa, around 5,600 years ago, which would suggest that my paternal ancestors spent over 4,000 years living in the Sahara. M183 is sometimes referred to as the Berber marker, since in some groups of Berbers, nearly 100% of the population exhibit this mutation. The Moors who invaded Spain were largely of Berber ethnicity.
Generations 186-750: From the Nile to the Sahara
Haplogroup E1b1b (M215), ancestral to E1b1b1b1b (M183) is believed to have emerged in eastern Africa, around 22,400 years ago, perhaps along the Nile in present-day Ethiopia or Sudan. 750 generations ago, my direct paternal ancestor may have spoken an ancient language, ancestral to the modern Nilo-Saharan languages.
Generations 751-1200: From the Ethiopian Highlands
Haplogroup E1b1 (P2/PN2), ancestral to E1b1b (M215) is believed to have emerged in the Ethiopian highlands, around 35,000 years ago. 1200 generations ago, my direct paternal ancestor may have spoken a language not only ancestral to the Nilo-Saharan languages, but also to the Niger-Congo and Mande languages.
Tuesday, October 11, 2011
The Maasai of Kenya
 The Maasai are an ethnic group of semi-nomadic people located in Kenya and northern Tanzania. They are among the best known of African ethnic groups, due to their distinctive customs and dress and residence near the many game parks of East Africa. I came across these four Maasai tribesmen in Nairobi National Park, Keyna.
The Maasai are an ethnic group of semi-nomadic people located in Kenya and northern Tanzania. They are among the best known of African ethnic groups, due to their distinctive customs and dress and residence near the many game parks of East Africa. I came across these four Maasai tribesmen in Nairobi National Park, Keyna.
Maasai DNA
Recent studies of Maasai DNA reveal a complex history of the Maasai people. In 2005, Elizabeth Wood, et al., sampled the Y-DNA of 26 Maasai tribesmen, and from this sample determined that 50% of the individuals had Y-DNA of Haplogroup E1b1b1 (M35), 27% had A3b2 (M13), 16% had E1b1a1 (M2), and 8% B2a (M150). An explanation of each, in the context of the history of the Maasai, follows.
Eastern Sudanic Ancestry
The largest Y-DNA found among the Maasai sample, E1b1b1, represents Eastern Sudanic ancestry (although it is also associated with certain other "Nilo-Saharan" populations). One would expect to see a high percentage of this haplogroup based on the fact that the Maasai speak a Nilotic (Eastern Sudanic) language. However, based on the fact that only 50% of their DNA is Eastern Sudanic, it would appear that the Maasai are remnants of an older culture whose language went extinct after prolonged contact and interbreeding with Nilotic peoples. Examining the other components of their Y-DNA is the key to understanding the origin of the Maasai.
Niger-Congo Ancestry
Haplogroup E1b1a1 represents a component of the Masaai ancestry common among most Sub-Saharan populations, indicating Niger-Congo ancestry. The most likely source of this haplogroup is the Bantu expansion, whereby Bantu-speaking peoples spread across Sub-Saharan Africa from East to West, generally. However, this haplogroup does not reveal the ancient origin of the Masaai people.
Pygmy and Khoisan Ancestry
It would appear that prior to the Bantu expansion and the southward migration of the Nilotic peoples, the Maasai were a combination of Pygmy and Khoisan people, assuming that haplogroup B is the African Pygmy modal haplogroup and A3b is the Khoisan or "Bushman" modal haplogroup. The deep components of Masaai DNA suggest an admixture between the most divergent branch of the Pygmies and the most divergent branch of the Khoisan. Most Pygmy populations exhibit a significant percent of the B2b clade, whereas the Maasai exhibit the B2a sister clade. Most Khoisan peoples exhibit a significant percent of the A3b1 clade, whereas the Maasai exhibit the A3b2 sister clade. It would appear that the distant ancestors of the early Maasai (excluding recent Supra-Saharan admixture) were a mixture of members of two ancient African hunter-gatherer cultures. The spread of Nilotic and Bantu culture wiped out the entire language families of the component populations, although the Maasai maintained much of their ancient nomadic culture.
Maasai DNA Reveals Origins of Sandawe People
Until recently, the Sandawe people, although living in Tanzania, were considered to be closely related to the Khoisan ethnicities of the Kalahari desert. Much of this presumption was based on the fact that like the Kalahari Bushmen, the Sandawe speak a click language. However, in 2007, Sarah Tishkoff, et al., conducted a study on the Y-DNA of 68 Sandawe people. The results were more complicated than even the Maasai. However, excluding the 56 individuals who tested for Supra-Saharan Y-DNA (Eastern Sudanic and Niger-Congo haplogroups described above), the remaining 12 individuals exhibited 72% haplogroup B2b (M112), 22% haplogroup A3b2 (M13) and 6% B2a (M150). Note the 22:6 ratio of A3b2 to B2a, comprared with the nearly identical 27:8 ratio among the Maasai. This suggests that the Sandawe were originally B2b pygmies, with subsequent admixtures from (not necessarily in this order) the Maasai, Bantus and Nilotics. Unlike the Maasai, however, the Sandawe retained their ancient click language. Since the Hadza people of Tanzania are the nearest B2b click-speaking tribe, one could presume that the language of the Sandawe is distantly related to the Hadza language, both languages perhaps descending from a common language spoken by the original Pygmy who underwent the B2b Y-DNA mutation. That would mean that the only extant "true" A3b1 Khoisan languages are those click-languages spoken in and around the Kalahari desert.
Implications Beyond Sub-Saharan Africa
According to the most recently accepted version of the mt-DNA phylogenetic tree, it is believed that the first split occurred separating the Khoisan L0 clade from the L1-6 superclade (representing the founding populations of the speakers of all non-Khoisan languages). In the non-Khoisan superclade (represented by haplogroup BT in the Y-DNA phylogenetic tree), the first node appears to separate the African Pygmies from the ancestor of the speakers of all non-Khoisan and non-pygmy languages. This split corresponds to the split between B and CT in the Y-DNA tree. It is interesting that both the haplogroup A Khoisan languages and the haplogroup B Pygmy languages are click languages, and that CT (which is downstream from BT) is the only clade originating in this time period not dominated by click languages. This is some evidence, although not conclusive, that there may have been at one time a Proto-World language that had clicks among its sound inventory, ancestral to all modern languages, including the CT languages (including Enlgish, for example). That is to say, perhaps around 75,000-100,000 years ago, all languages had clicks, and the Supra-Saharan CT branch lost its clicks. An analogy would be the English language having lost grammatical gender despite its Indo-European origin.
References
History of Click-Speaking Populations of Africa Inferred from mtDNA and Y Chromosome Genetic Variation. Tishkoff, Sarah A. et al 2007.
Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes. Wood, Elizabeth T et al 2005.
Sunday, October 2, 2011
Basque Y-DNA
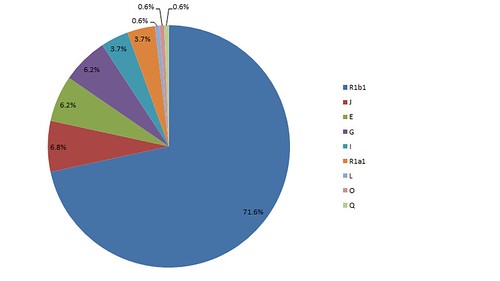
The above chart includes data from 162 male volunteers who submitted their Y-chromosomal DNA results to Family Tree DNA's Basque DNA project. Individuals who submitted their Y-DNA results claim to be of direct male Basque descent. Contributing volunteers included residents of Europe, Asia, and North and South America. Analysis of the data reveals that 71.6% of the participants in this study carry Y-DNA of haplogroup R1b1 and its subclades.
Basque People
The Basques as an ethnic group, primarily inhabit an area traditionally known as the Basque Country, a region that is located around the western end of the Pyrenees on the coast of the Bay of Biscay and straddles parts of north-central Spain and south-western France. Since the Basque language is unrelated to Indo-European, it is often thought that they represent the people or culture who occupied Europe before the spread of Indo-European languages there.
Y-DNA in the Field of Linguistics
Y-DNA haplogroup testing is a valuable tool in the study of historical linguistics. Y-DNA is carried from father to son, and mutates at a somewhat predictable rate. Haplogroups are clades of DNA types that share a distinct defining mutation or mutations. Each such mutation occurred in a single person at some point in the past. Since the person in which that mutation occurred necessarily spoke a language (at least in the time-frame of the past 50,000 years or so), and since a large percentage of children learn to speak the same native language as their father, one can use Y-DNA studies to track the historical evolution of languages, and uncover ancient relationships between living language families, to a surprising degree of accuracy.
When using Y-DNA as a tool to discover relationships between living language families, however, one must take into account the fact that there are several reasons why children may not learn to speak the native language of their fathers. The most obvious of such a situation is when the father either moves to a region that speaks a different language or has a child in a region where another language is dominant in addition to his native language (i.e., a more dominant local language is taught in schools, used in business, etc.), and rather than learning the native language of the father, children adopt the local language.
The goal in interpreting the data from this study is to determine which, if any, of the individuals whose Y-DNA first contained the defining mutations of the haplogroups, may have spoken a language ancestral to modern Basque, i.e. "Ancient Basque."
Haplogroup E Among Basque People
93.8% of those tested reported haplogroups of Eurasian origin, whereas 6.2% reported haplogroup E and its subclades. Haplogroup E is common among ethnic groups which originated along northern portions of the Nile River in Africa, including speakers of Nilo-Saharanm, Niger-Congo, Mande and certain Afro-Asiatic languages. The infusion of Y-DNA haplogroup E among the Basque population likely took place long after speakers of the ancestral Basque language arrived on the Iberian Peninsula, perhaps after the Afro-Asiatic speaking Moors invaded southern Europe. That is to say, the Basque language is unlikely to share a common origin with the Nilo-Saharan or Afro-Asiatic language families, although males of Northern African descent who migrated north to the Iberian Peninsula apparently interbred with women of the Basque population, perhaps influencing the local "Ancient Basque" language, but not replacing it.
Outliers in Haplogroups L, O and Q
Of the 162 individuals tested, there was one individual who carried haplogroup L, one who carried haplogroup O, and one who carried haplogroup Q. These haplogroups are of Eurasian origin, but are probably not associated with speakers of Ancient Basque. The individual who carried haplogroup Q resided in China and reported that his most distant known paternal ancestor resided in Mexico and had a Basque surname. It should be noted that Y-DNA haplogroup Q is the most common haplogroup among native (non-European) Mexicans, and not likely indicative of a Basque origin, despite the Basque surname. Haplogroups L and O generally correspond with South Asian and Asiatic languages, respectively. As the present-day Basque language shares little in common with members of these well-studied language families, the individuals likely represent a very small segment of the Basque population who descend from recent (less than 5000 years ago) immigrants to the Basque country from southern and eastern Asia.
Haplogroup R1a1 on the Iberian Peninsula
While haplogroup R1a1 appears in this sample at a percentage of 3.7%, that rate is similar to, if not less than, the occurrence of R1a1 in surrounding regions. R1a1 likely corresponds to DNA of native speakers of Indo-European languages, who settled the Iberian Peninsula and likely wiped out all recent branches of the Ancient Basque language with the exception of the languages of the Basque country. Haplogroup R1a1 is found in all locations where Indo-European languages are spoken, and the person in which its defining mutation occurred likely spoke a language ancestral to Indo-European, not to Basque.
Northwest Caucasian Haplogroup G
Y-DNA haplogroup G has not been definitively associated with any living language family, although it is common among speakers of Northwest Caucasian languages. The language of the progenitor of haplogroup G may only be manifested in the Northwest Caucasian substrate which differentiates the Northwest from the Northeast Caucasian languages. Since there are unlikely any modern surviving languages that descend directly from the language spoken by the progenitor of haplogroup G, it is difficult to rule out the haplogroup as corresponding to an Ancient Basque precursor. However, due to the overwhelming majority of haplogroup R1b1 (which shares the same lack of known modern surviving descendant languages) among the Basque population, it seems logical that many carriers of haplogroup G may have spoken a Vasconian (pre-Basque) language (perhaps since as long as 10,000 years ago), after native Vasconian speaking carriers of R1b1 dominated their native culture, perhaps shortly after the last ice age.
Cro-Magnon Haplogroup IJ
10.5% of the sample reported haplogroups of either I or J, both haplogroups that represent subsequent mutations from an earlier Cro-Magnon haplogroup IJ, which appears to have originated in the Caucasus. This is a large percentage of the sample that cannot be discounted. The progenitor of haplogroup J may have spoken a language ancestral to the Northeast Caucasian and Kartvelian language families. The progenitor of haplogroup I spoke a language belonging to an extinct family that may only have modern observable manifestation in the substrate of vocabulary found in the Germanic languages (approximately 1/3 of the lexicon) that is not traceable to Proto-Indo-European origin. While neither haplogroups I nor J can be definitively ruled out as corresponding to the Basque language, it appears that the Basque language does not share much in common with the Northeast Caucasian or Kartvelian languages, nor have I found any source that suggesting that its lexicon overlap the Proto-Germanic substrate.
Vasconian Haplogroup R1b1
Based on the data from this study in a vacuum, it seems very likely that if the progenitors of any of these haplogroups spoke a Vasconian language, it should be R1b1, as R1b1 accounts for 71.6% of the sample population. However, looking outside this study, R1b1 is equally common among most Indo-European speaking (Spanish and Portuguese, e.g.) populations of the Iberian peninsula, and almost as common in regions to the east where other Romance languages are spoken such as French and Italian. Perhaps remnants of the language spoken by the progenitor of R1b1 can be found by studying the differences between the Romance (Italic) brancih of the Indo-European languages from other Indo-European subfamilies. I suspect some of the differences may be accounted for by a Vasconian substrate that represents linguistic elements of other languages descended from Vasconian that may have been spoken by populations who assimilated into the western Indo-European culture, and adpoted Indo-European as their language. I believe this hypothesis is more sound than a IJ origin of the Basque language, based on geographic data on the present location of haplogroup R1b1 vs. haplogroups I and J. For example, haplogroup I is distributed widely in Scandinavia and in regions where Germanic languages are spoken. It seems likely that if there was a living descendant language of the language spoken by the progenitor of haplogroup I, it would have the highest likelihood of surviving in Germanic speaking Europe, not in the Pyrenees Mountains where R1b1 y-DNA is dominant among Italic speakers who (if they inherited their language from their ancestors rather than by assimilation) would be expected to have R1a1 Indo-European DNA.
Basque Language Isolate
One might ask, if the Basque language is associated with R1b1, and the Indo-European languages are associated with R1a1, (both clades of R1), why is the modern Basque language so different from all of its closest genetic relatives? However, consider, for example, the incredible difference between the English and Hindi languages (both Indo-European) which probably only diverged from their most common ancestor about 5,000 years ago. If not for available linguistic data from the numerous other languages in the Indo-European family, one might be highly skeptical about their common origin, especially due to the geographic distance where the two languages are spoken, and the differences in culture, appearance, religions, etc., between the populations by whom they are spoken. One must keep in mind that R1a and R1b diverged from their common y-DNA ancestor R1 approximately 18,000 years ago, and there are no intermediate languages on the R1b side that survived to modern times, with the possible exception of Basque. It should not be surprising that Basque seems completely foreign to the Indo-European languages, in this context.
Furthermore, even if the progenitor of R1a spoke an ancient Indo-European language and the progenitor of R1b spoke an ancient Vasconian language, that does not necessarily imply that the two ancient languages were closely related. The progenitor of R1 (ancestral to R1a and R1b) probably lived in Siberia some 25,000 to 30,000 years ago, approximately 10,000 years prior to the mutations that occurred that created the subclades of R1a and R1b. In those 10,000 years, the descendants of the progenitor of R1 may have come to speak many languages unrelated to the the native language of their ancestor by means of assimilation as they migrated across Asia. That is to say, while R1a and R1b carriers are undoubtedly genetically related, an inter-disciplinary approach including efforts by experts in the fields of anthropology, archaeology, linguistics, genetics, philology and other sciences, is required to prove or disprove any ancient relation between the Basque language and the Indo-European languages. Also, an examination of the Burushoski language spoken by (among others) descendants of R1's closest relative R2 may provide some evidence helpful to determining the origin of the Basque language. Unfortunately, Burushoski, spoken in portions of present-day Pakistan, is also a language isolate.
Related Reading
For what they were... we are: Linguistic musings: Basque and Proto-Indoeuropean
Diagram, research and analysis by Kevin Borland. Data provided by Family Tree DNA. Text of subsection "Basque People" derived from Wikipedia.
Sunday, September 25, 2011
Roger Clyne at the State Theatre

Roger Clyne and the Peacemakers performed September 25, 2011, at the State Theatre in Falls Church, Virginia. They played the following songs:
1. All Over the Radio
2. Down Together
3. Heaven on a Paper Plate
4. Banditos
5. Dinero
6. Mexico
7. Lemons
8. Marie
9. Tell Yer Momma
10. Maybe We Should Fall in Love
11. Interstate
12. Paper Airplanes
13. Wanted
14. I Know You Know
15. Girly
16. Yahoos and Triangles
17. Empty Highway
18. European Swallow
19. Mekong
20. Green and Dumb (encore)
21. Counterclockwise (encore)
22. Nada (encore)
Photo and videos by Kevin Borland.
Saturday, September 24, 2011
Trip to the Zoo
I took my mother to visit the National Zoo today. I have visited the few a handful of times before, but this time I spent quite a bit of time in the Small Mammal House. Here are a few of the animals we saw:
White-Faced Saki
The white-faced saki, also known as the Guianan saki and the golden-faced saki, is a species of saki monkey, a type of New World monkey, found in Brazil, French Guiana, Guyana, Suriname, and Venezuela. This species lives in the understory and lower canopy of the forest, feeding mostly on fruits, but also eating nuts, seeds, and insects.
Golden Lion Tamarin
The golden lion tamarin is diurnal and primarily arboreal, forming small groups of patrols with a consistent territory of around 400,000 square metres (100 acres). Fighting between groups is avoided by scent marking and ritual encounters. By night, the tamarins sleep in abandoned nesting holes in trees, or in large bromeliads.


Pale-faced saki (left), golden lion tamarin (right). Photos by Kevin Borland. Portions of text derived from Wikipedia.
White-Faced Saki
The white-faced saki, also known as the Guianan saki and the golden-faced saki, is a species of saki monkey, a type of New World monkey, found in Brazil, French Guiana, Guyana, Suriname, and Venezuela. This species lives in the understory and lower canopy of the forest, feeding mostly on fruits, but also eating nuts, seeds, and insects.
Golden Lion Tamarin
The golden lion tamarin is diurnal and primarily arboreal, forming small groups of patrols with a consistent territory of around 400,000 square metres (100 acres). Fighting between groups is avoided by scent marking and ritual encounters. By night, the tamarins sleep in abandoned nesting holes in trees, or in large bromeliads.
Pale-faced saki (left), golden lion tamarin (right). Photos by Kevin Borland. Portions of text derived from Wikipedia.
Sunday, September 18, 2011
Blacksmith at Gulf Branch Forge

I wanted to get out of the house today, so I went over to the Gulf Branch Forge, where I met this friendly blacksmith. I watched her make an ornate hook out of a steel rod. First she made each end of the rod pointed and rounded using the hammer and anvil. Then, she bent one of the ends into a spiral for decoration. Next, she formed the hook portion by wrapping the bar around a circular piece of metal. Then, she bent the straight end at a 90 degree angle so it could be hammered into a wall. She then proceeded to twist the middle of the hook (between the 90 degree angle an the curved portion) for a decorative effect. Finally, she straightened it and polished it. The piece was finished about a half hour from when she first placed the rod in the forge. While she was making the hook she explained every step of the process, and discussed everything from how the coal forge works to how lasers are used to check the insulation on modern gas forges.
Photo by Kevin Borland.
Subscribe to:
Posts (Atom)